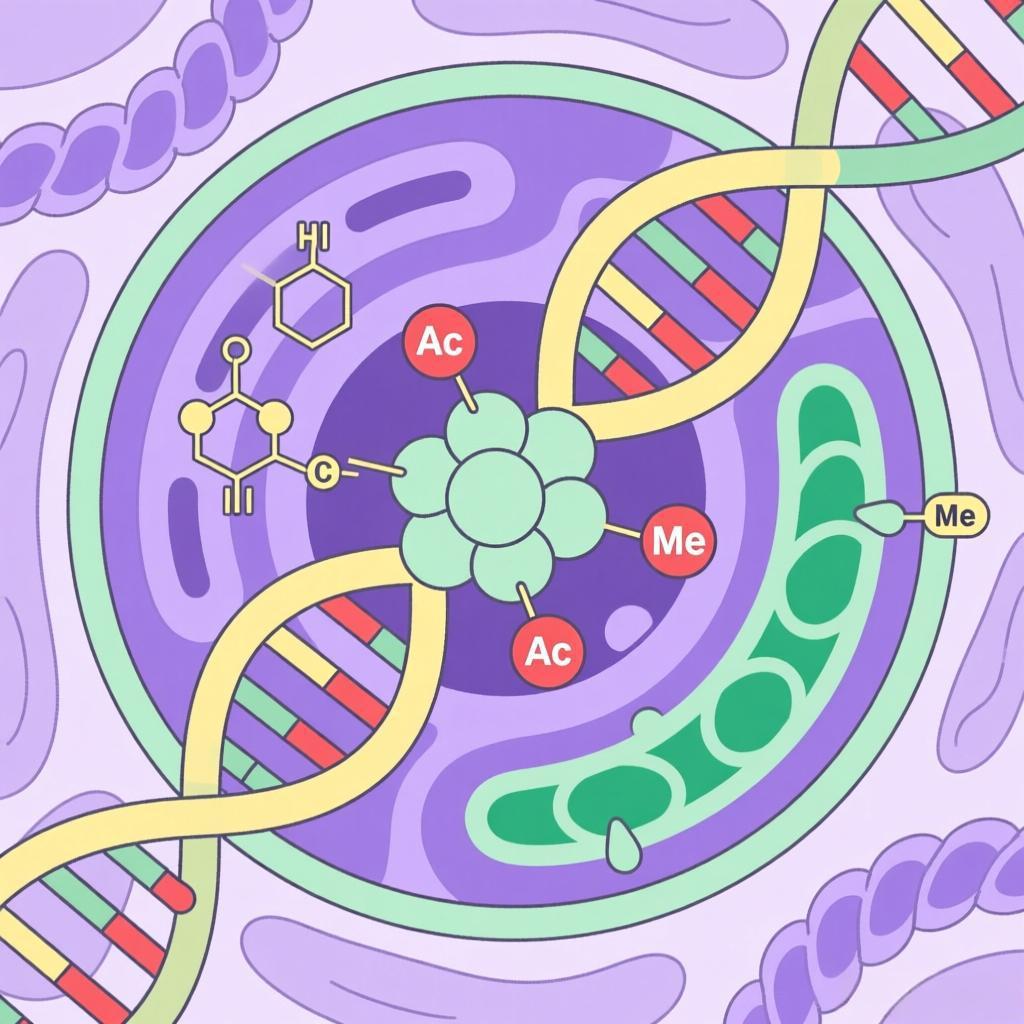
表观遗传机制
表观遗传机制是连接基因型与表型的关键,不改变DNA序列却能调控基因表达,核心包括DNA甲基化、组蛋白修饰等四大类型,具有可逆性、可遗传性、环境响应性。
中文名:
表观遗传机制英文名:
Epigenetic Mechanism核心定义:
不改变DNA序列,通过化学修饰等方式调控基因表达,且修饰状态可通过细胞分裂遗传的分子机制核心类型:
DNA甲基化、组蛋白修饰、非编码RNA调控、染色质重塑本质特征:
可逆性、可遗传性、环境响应性、不改变DNA序列核心意义:
调控细胞分化、参与疾病发生、介导环境与基因的相互作用机制类型
一、DNA甲基化
DNA甲基化是研究最深入、最具代表性的表观遗传机制之一,也是最早被发现的表观遗传修饰方式,主要发生在DNA分子中胞嘧啶(C)的5位碳原子上,通过添加一个甲基(-CH₃)形成5-甲基胞嘧啶,这一过程由DNA甲基转移酶(DNMTs)催化完成,而甲基的去除则由去甲基化酶(如TET家族蛋白)介导,具有动态可逆的特点。在真核生物中,DNA甲基化主要集中在基因组中的CpG二核苷酸序列上(即胞嘧啶与鸟嘌呤通过磷酸二酯键相连的序列),这些序列往往成簇存在,形成“CpG岛”,多位于基因的启动子区域。当CpG岛发生高度甲基化时,会阻碍转录因子与启动子区域的结合,同时招募具有抑制作用的染色质结合蛋白,从而抑制基因的转录表达,实现基因沉默;而CpG岛去甲基化则会解除这种抑制,使基因重新被激活。DNA甲基化的核心功能的包括三个方面:一是参与细胞分化调控,在胚胎发育过程中,不同细胞类型会形成特异性的DNA甲基化模式,锁定细胞的分化状态,确保肝细胞、神经细胞、肌肉细胞等不同细胞只表达自身功能所需的基因;二是维持基因组稳定性,通过沉默转座子、病毒序列等外源或异常DNA,防止其插入正常基因组导致基因突变;三是参与基因组印记和X染色体失活,例如女性体细胞中一条X染色体通过广泛甲基化实现沉默,避免X染色体上基因的过度表达,而基因组印记则通过亲代特异性的甲基化模式,使父源和母源基因呈现差异性表达,确保胚胎正常发育。需要注意的是,DNA甲基化模式会受到环境因素的显著影响,年龄增长、饮食不均衡、吸烟、长期压力、化学毒素暴露等,都可能导致DNA甲基化异常。这种异常与多种疾病密切相关,例如在癌症中,肿瘤抑制基因启动子区域的高甲基化会导致其沉默,而癌基因的低甲基化则会使其异常激活,共同推动肿瘤的发生和发展;在神经退行性疾病中,DNA甲基化异常会影响神经细胞相关基因的表达,导致神经功能损伤。
")
二、组蛋白修饰
真核生物的DNA并非裸露存在,而是缠绕在由组蛋白组成的核心颗粒上,形成核小体,核小体进一步折叠、压缩形成染色质。组蛋白是一类碱性蛋白质,主要包括H2A、H2B、H3、H4四种核心组蛋白和H1一种连接组蛋白,其中核心组蛋白的N端尾巴会伸出核小体表面,可发生多种翻译后修饰,这些修饰被称为组蛋白修饰,是表观遗传机制的另一核心类型。目前已发现的组蛋白修饰类型包括乙酰化、甲基化、磷酸化、泛素化、 SUMO化等,不同类型的修饰以及修饰位点、修饰程度,都会对染色质结构和基因表达产生不同的影响,这些修饰的组合形成了复杂的“组蛋白密码”,协同调控基因表达。其中,乙酰化和甲基化是最常见、研究最深入的两种修饰。组蛋白乙酰化主要由组蛋白乙酰转移酶(HATs)催化,将乙酰基(-COCH₃)添加到组蛋白的赖氨酸残基上;去乙酰化则由组蛋白去乙酰化酶(HDACs)介导,去除乙酰基。乙酰化会中和组蛋白的正电荷,降低组蛋白与带负电DNA的结合力,使染色质结构变得松散(形成常染色质),便于转录因子和RNA聚合酶与DNA结合,从而促进基因表达;而去乙酰化则会增强组蛋白与DNA的结合,使染色质压缩(形成异染色质),抑制基因表达。组蛋白甲基化主要发生在组蛋白的赖氨酸和精氨酸残基上,由组蛋白甲基转移酶(HMTs)催化,去甲基化则由组蛋白去甲基化酶(HDMs)介导。与乙酰化不同,组蛋白甲基化的功能具有多样性,取决于修饰的位点和甲基化程度:例如,组蛋白H3第4位赖氨酸的三甲基化(H3K4me3)通常与基因启动子激活相关,而H3第9位赖氨酸的三甲基化(H3K9me3)和H3第27位赖氨酸的三甲基化(H3K27me3)则与基因沉默相关,参与异染色质的形成和维持。组蛋白修饰的核心作用是通过改变染色质的空间结构,调控基因的可及性,同时招募不同的调控蛋白,参与基因表达的精细调控。其异常同样与疾病密切相关,例如,HDACs过度激活会导致组蛋白去乙酰化异常,抑制肿瘤抑制基因表达,促进癌症发生;而组蛋白甲基转移酶NSD2的异常激活,会导致癌症细胞发生“谱系可塑性”,增强其耐药性和侵袭性。此外,组蛋白修饰异常还与神经退行性疾病、免疫失调等疾病相关。三、非编码RNA调控
非编码RNA(ncRNA)是一类不编码蛋白质,但具有明确调控功能的RNA分子,其长度从几十个核苷酸到几千个核苷酸不等,根据长度可分为微小RNA(miRNA,18-25nt)、长链非编码RNA(lncRNA,>200nt)、PIWI互作RNA(piRNA)等。非编码RNA通过多种方式参与表观遗传调控,是连接DNA修饰、组蛋白修饰与基因表达的重要桥梁,也是近年来表观遗传研究的热点领域。微小RNA(miRNA)是最常见的非编码RNA类型之一,其调控机制主要是通过碱基互补配对,特异性结合靶标mRNA的3'非翻译区(UTR),导致靶标mRNA降解或抑制其翻译,从而在转录后水平沉默基因表达。一个miRNA可以调控多个靶标mRNA,而一个mRNA也可以被多个miRNA调控,形成复杂的调控网络,参与细胞分化、凋亡、代谢等多种生命活动。例如,miR-21可通过沉默肿瘤抑制基因,促进癌症的发生和发展;而miRNA的异常表达还与心血管疾病、神经发育异常等相关。长链非编码RNA(lncRNA)的调控方式更为多样,主要包括四个方面:一是作为“分子支架”,招募组蛋白修饰酶、DNA甲基转移酶等调控因子,靶向结合到特定基因组位点,调控该区域的DNA甲基化和组蛋白修饰,从而影响基因表达;二是作为“分子海绵”,吸附miRNA,阻止其与靶标mRNA结合,解除miRNA对基因的沉默作用;三是直接与DNA或RNA结合,改变其空间结构,影响基因转录或mRNA剪切;四是参与染色质重塑,调控染色质的开放与关闭状态。例如,lncRNA XIST可招募组蛋白修饰酶,介导女性体细胞中一条X染色体的沉默,参与X染色体失活过程;而部分lncRNA的异常表达会导致癌症、神经退行性疾病等。PIWI互作RNA(piRNA)主要存在于生殖细胞中,其核心功能是沉默转座子,维持生殖细胞基因组的稳定性,同时参与生殖细胞的发育和成熟。piRNA通过与PIWI蛋白结合形成复合物,识别并降解转座子RNA,或诱导转座子区域发生DNA甲基化和组蛋白修饰,从而抑制转座子的活性,避免其对生殖细胞基因组造成损伤。此外,近年来研究发现,精子中的miRNA等非编码RNA还可作为表观遗传信息载体,介导父代运动能力、代谢状态等性状的代际遗传。四、染色质重塑
染色质重塑是指在染色质重塑复合物的作用下,通过改变核小体的位置、组成或结构,实现染色质结构动态变化的过程,其核心是利用ATP水解提供的能量,调控染色质的开放(常染色质)与关闭(异染色质)状态,从而影响基因的可及性和表达水平。染色质重塑与DNA甲基化、组蛋白修饰、非编码RNA调控密切协同,共同构成表观遗传调控网络。染色质重塑复合物主要分为SWI/SNF、ISWI、CHD、INO80等多个家族,不同家族的复合物具有不同的功能:例如,SWI/SNF家族复合物可通过移动核小体的位置,使基因启动子区域暴露,促进转录因子结合,从而激活基因表达;而ISWI家族复合物则主要参与核小体的组装和排列,维持染色质的稳定结构。染色质重塑的核心功能是参与细胞分化、胚胎发育、基因组稳定性维持等过程。在胚胎发育过程中,染色质重塑复合物通过调控不同基因区域的染色质状态,推动细胞分化为不同的组织和器官;在细胞分裂过程中,染色质重塑参与染色质的凝聚和分离,确保染色体的正确传递;此外,染色质重塑还可响应环境信号,通过改变染色质结构,调整基因表达模式,帮助细胞适应环境变化。染色质重塑异常同样与疾病密切相关,其中最典型的是癌症:例如,SWI/SNF复合体成分ARID1A的突变在多种恶性肿瘤中频繁出现,导致染色质重塑功能紊乱,基因表达异常,推动肿瘤发生;而染色质重塑异常还与神经发育疾病、免疫缺陷等疾病相关。华东师范大学翁杰敏教授团队与中国科学院相关团队合作,首次揭示了哺乳动物细胞中控制染色质分区以及近着丝粒异染色质形成、维持和稳态遗传的新机制,发现泛素连接酶G2E3可通过催化组蛋白H3第14位赖氨酸的单泛素化修饰,调控异染色质的稳态遗传,这一发现为理解染色质遗传机制提供了新视角。核心特征
一、不改变DNA序列
这是表观遗传机制与经典遗传学最本质的区别。经典遗传学通过DNA序列的改变(如碱基替换、缺失、插入)来改变遗传信息,进而影响性状;而表观遗传机制仅通过DNA甲基化、组蛋白修饰等化学修饰,或染色质结构的改变,调控基因表达,不涉及DNA碱基序列的任何变化。例如,同卵双胞胎具有完全相同的DNA序列,但由于表观遗传修饰的差异,可能会在身高、性格、疾病易感性等方面出现差异,这正是表观遗传机制作用的结果。二、可逆性
与DNA突变的不可逆性不同,表观遗传修饰具有动态可逆的特点。DNA甲基化可以通过去甲基化酶的作用被去除,组蛋白修饰可以通过去修饰酶的作用恢复到未修饰状态,非编码RNA的表达也可以被调控,染色质结构也可以在重塑复合物的作用下发生动态变化。这种可逆性使得细胞能够根据外界环境的变化,灵活调整基因表达模式,适应环境需求,同时也为疾病的治疗提供了可能——通过调控表观遗传修饰,恢复异常基因的正常表达,从而达到治疗疾病的目的。例如,HDAC抑制剂可以抑制组蛋白去乙酰化酶的活性,恢复肿瘤抑制基因的表达,已被用于癌症的临床治疗。三、可遗传性
表观遗传修饰具有可遗传性,主要分为细胞水平的遗传和跨代遗传两种形式。细胞水平的遗传是指在细胞分裂过程中,亲代细胞的表观遗传修饰模式会被精确传递给子代细胞,确保子代细胞维持与亲代细胞相同的基因表达模式和细胞功能。例如,肝细胞分裂后,子代肝细胞依然保持肝细胞的特异性功能,正是由于表观遗传修饰模式的稳定传递。跨代遗传则是指表观遗传修饰模式通过生殖细胞(精子、卵子)传递给后代,使后代在不改变DNA序列的情况下,继承亲代的某些表型或疾病易感性。例如,“荷兰饥饿冬季”研究显示,孕期经历饥荒的妇女,其后代出现了DNA甲基化模式的改变和代谢疾病风险的增加;中国科学家首次发现,精子miRNA可作为表观遗传信息载体,实现父代运动能力的代际遗传;而母亲高血糖则会干扰原始生殖细胞发育过程中的性别特异性表观遗传重编程,导致后代生育能力下降。需要注意的是,人类的跨代表观遗传机制仍在深入研究中,其具体规律和机制尚未完全明确。四、环境响应性
表观遗传机制具有很强的环境响应性,外界环境因素(如饮食、压力、吸烟、饮酒、化学毒素暴露、温度变化、缺氧等)可以通过多种途径诱发表观遗传修饰的改变,进而调整基因表达模式,影响个体的健康状态和疾病易感性。这种响应性使得生物能够更好地适应环境变化,是生物进化过程中形成的重要适应机制。例如,长期高糖饮食会导致体内表观遗传修饰异常,增加肥胖、糖尿病等代谢性疾病的风险;长期吸烟会改变肺部细胞的DNA甲基化和组蛋白修饰模式,增加肺癌的发病风险;缺氧环境会通过抑制组蛋白去甲基化酶KDM6A活性,以缺氧诱导因子(HIF)非依赖的方式抑制铁死亡,帮助细胞适应缺氧环境;而饮食中的营养物质(如叶酸、维生素B12)则参与DNA甲基化的合成,缺乏这些营养物质会导致DNA甲基化异常,影响基因表达。
")
生物学意义
一、调控细胞分化与胚胎发育
在胚胎发育过程中,受精卵从一个全能细胞逐渐分化为各种具有特异性功能的细胞(如神经细胞、肌肉细胞、肝细胞等),这一过程的核心是表观遗传机制的调控。在胚胎发育的不同阶段,不同的表观遗传修饰模式被建立,沉默与细胞特异性功能无关的基因,激活与细胞功能相关的基因,从而锁定细胞的分化方向,确保胚胎正常发育为完整的个体。例如,在胚胎干细胞中,染色质处于开放状态,大量与细胞全能性相关的基因被激活,而与细胞分化相关的基因被沉默;随着胚胎发育的推进,表观遗传修饰发生动态变化,与特定细胞类型相关的基因被激活,全能性基因被沉默,从而实现细胞的分化。如果表观遗传机制异常,会导致胚胎发育异常,甚至引发胚胎致死。二、维持基因组稳定性
表观遗传机制通过多种方式维持基因组的稳定性,防止基因组损伤和基因突变。一方面,DNA甲基化和非编码RNA调控可以沉默转座子、病毒序列等外源或异常DNA,避免其插入正常基因组导致基因突变;另一方面,组蛋白修饰和染色质重塑可以维持染色质的稳定结构,防止染色体断裂、缺失、重排等畸变,确保细胞分裂过程中染色体的正确传递。例如,piRNA通过沉默生殖细胞中的转座子,维持生殖细胞基因组的稳定性,确保生殖细胞的正常发育和遗传信息的正确传递;而染色质重塑复合物则通过调控染色质结构,避免染色体在细胞分裂过程中发生异常分离,减少染色体畸变的发生。三、介导环境与基因的相互作用
表观遗传机制是环境因素与基因相互作用的重要媒介,它能够将外界环境信号转化为细胞内的基因表达调控信号,使生物能够适应环境变化。环境因素通过诱发表观遗传修饰的改变,调整基因表达模式,进而影响个体的表型和疾病易感性,这种相互作用贯穿于个体的整个生命过程,甚至可以通过跨代遗传影响后代的健康。例如,牛津大学等机构的研究人员发现,父亲的饮食、肥胖、吸烟、内分泌干扰物暴露及压力等因素,会改变精子中的DNA甲基化、组蛋白保留和小非编码RNA等表观遗传标记,进而影响后代的健康状态;而孕期母亲的营养状况、压力水平等,也会通过表观遗传机制影响胎儿的发育,增加后代患代谢性疾病、神经发育异常等疾病的风险。四、参与疾病的发生与发展
表观遗传机制的异常是多种疾病发生、发展的核心原因之一,几乎所有疾病(包括癌症、神经退行性疾病、代谢性疾病、免疫疾病等)都与表观遗传修饰异常相关。与基因突变相比,表观遗传修饰异常具有可逆性,这为疾病的诊断和治疗提供了全新的靶点,成为现代医学研究的重要方向。在癌症中,表观遗传异常主要表现为肿瘤抑制基因的高甲基化沉默、癌基因的低甲基化激活、组蛋白修饰异常以及非编码RNA表达紊乱,这些异常共同推动肿瘤细胞的增殖、侵袭、转移和耐药性;在神经退行性疾病(如阿尔茨海默病、帕金森病)中,表观遗传修饰异常会影响神经细胞的存活和功能,导致神经纤维缠结、神经元凋亡等病理变化;在代谢性疾病(如糖尿病、肥胖)中,表观遗传异常会影响代谢相关基因的表达,导致代谢紊乱。
")
深度解读
表观遗传机制的发现,彻底改变了人们对遗传规律的认知,打破了“DNA序列决定一切”的传统观念,揭示了生命调控的复杂性和多样性。从本质上来说,表观遗传机制是生命体内一套“动态的基因表达调控系统”,它既能够稳定传递遗传信息,确保细胞功能的稳定性和遗传性,又能够灵活响应环境变化,调整基因表达模式,使生物能够适应复杂的外界环境。从进化角度来看,表观遗传机制是生物在长期进化过程中形成的重要适应机制。在复杂多变的环境中,生物无法通过快速改变DNA序列来适应环境,而表观遗传机制的可逆性和环境响应性,使得生物能够在不改变遗传信息的前提下,快速调整表型,适应环境变化,这种能力对于生物的生存和繁衍具有重要意义。同时,跨代表观遗传机制的存在,使得亲代对环境的适应能够传递给后代,帮助后代更好地适应环境,加速生物的进化。从医学角度来看,表观遗传机制为疾病的诊断和治疗提供了全新的思路和靶点。
传统的疾病治疗主要针对基因序列异常,而表观遗传治疗则针对表观遗传修饰异常,通过调控DNA甲基化、组蛋白修饰、非编码RNA表达等,恢复异常基因的正常表达,从而达到治疗疾病的目的。目前,已有多种表观遗传药物(如DNMT抑制剂、HDAC抑制剂)被批准用于癌症等疾病的临床治疗,且取得了良好的治疗效果。此外,表观遗传标记(如DNA甲基化、非编码RNA)还可以作为疾病的诊断标志物,用于疾病的早期诊断、预后判断和疗效监测,为精准医疗提供了重要支撑。需要注意的是,目前人类对表观遗传机制的研究仍处于不断深入的阶段,还有许多问题尚未完全明确,例如,跨代表观遗传的具体机制、不同表观遗传机制之间的协同调控关系、环境因素诱发表观遗传异常的具体途径等。随着高通量测序、CRISPR介导的表观遗传编辑等技术的发展,人们对表观遗传机制的认知将不断深化,其在生命科学、医学、农业等领域的应用也将不断拓展。
此外,表观遗传机制还具有重要的社会意义。它揭示了环境因素对人类健康的深远影响,强调了健康生活方式(如合理饮食、规律作息、远离烟酒、缓解压力)的重要性——良好的生活方式可以维持正常的表观遗传修饰,减少疾病的发生风险;而不良的生活方式则会导致表观遗传异常,增加疾病的发病风险。同时,表观遗传机制也为遗传病的预防和干预提供了新的思路,通过调控表观遗传修饰,可以预防或缓解某些遗传病的症状,提高人类的健康水平。
")
研究消息
近年来,表观遗传机制领域的研究发展迅速,不断有新的发现和突破,尤其是在2025-2026年,多项重要研究成果相继发表,进一步深化了人们对表观遗传机制的认知,也为其临床应用提供了新的支撑,以下是部分最新研究消息:
1.2026年2月5日,《科学进展》(Sci Adv)发表西北大学严健团队的研究成果,该团队首次构建了一套整合性功能基因组学研究流程,在原发性与转移性结直肠癌细胞模型中,系统性解析了非编码SNP与CpG甲基化对增强子活性的调控网络,清晰揭示了结直肠癌发生、转移的全新分子机制,为结直肠癌的表观遗传治疗提供了新的靶点和思路。
2. 2025年11月30日,《自然》(Nature)发表研究论文,揭示了癌症细胞“谱系可塑性”的表观遗传调控机制。研究人员通过海量数据证实,组蛋白甲基转移酶NSD2在其中发挥核心作用,它通过调控组蛋白修饰,推动癌症细胞发生身份转换,增强其耐药性和侵袭性,同时研究还证实,这种“谱系可塑性”是可以被逆转的,为癌症耐药的治疗提供了新的方向。
3. 2025年10月17日,《自然》(Nature)发表华东师范大学翁杰敏教授团队与中国科学院生物化学与细胞生物学研究所陈德桂研究员团队的合作研究成果,首次揭示了哺乳动物细胞中控制染色质分区以及近着丝粒异染色质形成、维持和稳态遗传的新机制,发现泛素连接酶G2E3可通过催化组蛋白H3第14位赖氨酸的单泛素化修饰,调控异染色质的稳态遗传,这一发现改变了人们对异染色质调控机制的认知,也为理解多种疾病的发生机制提供了新视角。
4. 2025年10月13日,中国科学家在国际期刊发表研究成果,首次提供了精子miRNA作为表观遗传信息载体、可介导父代运动/代谢能力代际遗传的直接证据,该研究揭示了父代性状跨代传递的全新表观遗传机制,为研究跨代表观遗传提供了重要参考。
5. 2025年10月27日,牛津大学等机构的研究人员在国际期刊发表研究成果,综合多项证据指出,父亲的饮食、肥胖、吸烟、内分泌干扰物暴露及压力等因素,会改变精子中的DNA甲基化、组蛋白保留和小非编码RNA等表观遗传标记,进而影响后代的健康状态,为预防跨代健康风险提供了理论依据。
")
6. 2025年9月30日,《自然·化学生物学》(Nat Chem Biol)发表研究成果,研究人员开发出针对关键表观遗传调控因子的首创抑制剂,该抑制剂能够精确解码特定HAT复合物的功能,揭示了ATAC在维持癌症基因表达程序中的独特作用,也为开发用于人类疾病的其他复合物特异性表观遗传药物提供了新的见解。
7. 2025年7月29日,《细胞》子刊发表姜学军团队的研究成果,首次发现缺氧通过抑制组蛋白去甲基化酶KDM6A活性,以缺氧诱导因子(HIF)非依赖的方式抑制铁死亡,该研究揭示了缺氧环境下表观遗传调控的新机制,为缺氧相关疾病(如心肌梗死、脑梗死、癌症)的治疗提供了新的靶点。[1][2][3]
词条图片