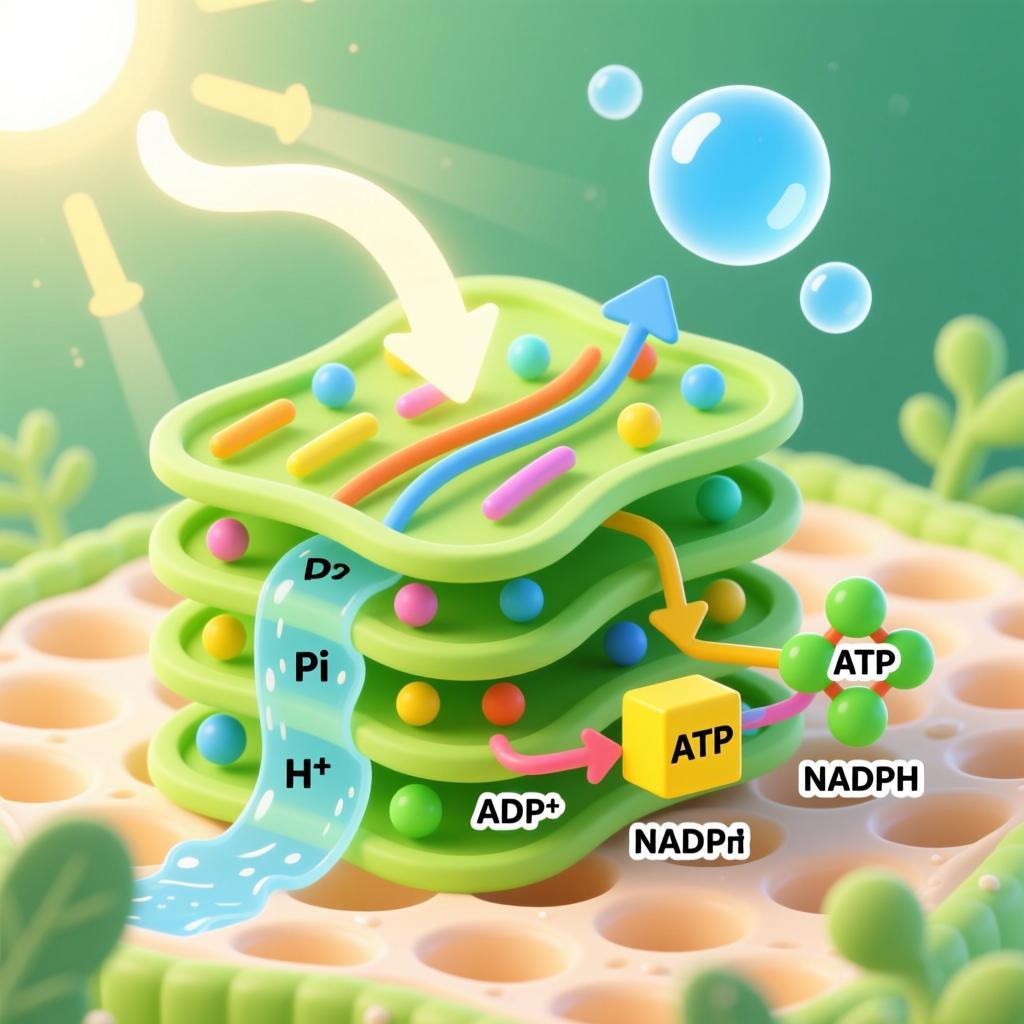
光合作用光反应阶段
光合作用光反应阶段是光合作用的核心步骤,本文详细介绍光反应的定义、反应场所、核心物质、三大关键过程,解析光反应与暗反应的区别与联系,说明其生理意义、应用价值,附最新研究进展,全面解答光反应相关疑问。
中文名称:
光合作用光反应阶段英文名称:
Light reaction stage of photosynthesis反应场所:
真核生物:叶绿体类囊体薄膜;原核生物(如蓝细菌):细胞质膜核心条件:
光照、光合色素、光反应酶、水能量转化:
光能→电能→活跃的化学能(储存在ATP和NADPH中)核心产物:
氧气(O₂)、三磷酸腺苷(ATP)、还原型辅酶Ⅱ(NADPH)反应基础:场所与物质准备
一、反应场所
光反应的场所具有高度特异性,不同类型的光合生物,其光反应场所存在差异,但均具备“膜结构”这一共同特征——膜结构可固定光合色素和酶系统,形成局部浓度优势,为电子传递和能量转化提供稳定环境。对于真核光合生物(如绿色植物、藻类),光反应的场所是叶绿体的类囊体薄膜。叶绿体是植物细胞中专门进行光合作用的细胞器,其内部由许多扁平的类囊体堆叠形成基粒,类囊体薄膜上分布着大量的光合色素、电子传递链蛋白和酶,是光反应的核心场所。类囊体内部形成类囊体腔,外部是叶绿体基质,这种膜结构的分隔的形成了质子梯度,为ATP的合成提供了必要条件。对于原核光合生物(如蓝细菌、光合细菌),由于其细胞内没有叶绿体等细胞器,光反应的场所是细胞质膜(又称细胞膜)。这类生物的细胞质膜上会折叠形成特殊的膜结构,其上镶嵌着光合色素和相关酶系统,模拟真核生物类囊体薄膜的功能,完成光能吸收和光反应过程。其中,蓝细菌的细胞质膜上还会形成类囊体样结构,进一步提高光反应的效率,其光反应过程与真核生物高度相似;而光合细菌(如紫硫细菌)的光反应则略有差异,不产生氧气,且电子供体为硫化氢而非水。二、核心物质:光合色素
光合色素是光反应中捕获光能的核心物质,主要分布在类囊体薄膜(或原核生物细胞质膜)上,与蛋白质结合形成色素蛋白复合体,分为叶绿素类、类胡萝卜素类两大类,不同色素的吸收光谱不同,共同作用可高效捕获可见光,提高光能利用率。叶绿素类是光合色素的主要类型,分为叶绿素a、叶绿素b(主要存在于绿色植物和藻类中),以及叶绿素c、叶绿素d(存在于部分藻类中)、细菌叶绿素(存在于光合细菌中)。其中,叶绿素a是唯一能参与光能转化的色素(即反应中心色素),其他叶绿素均为天线色素,仅负责吸收和传递光能,不能直接参与电子传递。叶绿素a的吸收峰主要在680nm(红光)和430nm(蓝紫光),叶绿素b的吸收峰在650nm(红光)和450nm(蓝紫光),二者协同作用,可覆盖可见光的主要波段,减少光能的浪费。类胡萝卜素类包括胡萝卜素和叶黄素,均为天线色素,主要吸收蓝紫光(吸收峰在400-500nm),其功能主要有两个:一是补充叶绿素对蓝紫光的吸收,提高光能捕获效率;二是保护叶绿素,防止其在强光下被氧化破坏(即光保护作用)。类胡萝卜素的颜色为橙黄色或黄色,当植物叶片衰老或环境胁迫时,叶绿素会分解,类胡萝卜素的颜色会显现出来,形成叶片变黄的现象。此外,部分藻类(如红藻、蓝细菌)还含有藻胆素(包括藻红素、藻蓝素),这类色素的吸收光谱与叶绿素差异较大,可吸收绿光和蓝光,适应海洋等弱光环境,进一步扩大了光合生物对光照的适应范围。所有光合色素均具有较多的共轭双键,可通过电子跃迁吸收光能,将光能转化为电子的势能,为后续的电子传递奠定基础。
")
三、其他关键物质
光反应的顺利进行,除了光合色素外,还需要水、光反应酶、电子传递链组分、ADP(二磷酸腺苷)、Pi(磷酸)、NADP⁺(辅酶Ⅱ)等物质的参与。其中,水是光反应的电子供体和原料,其分解产生的电子用于补充光合色素失去的电子,氢离子用于形成质子梯度和合成NADPH;光反应酶包括水裂解酶、ATP合成酶等,负责催化水的分解和ATP的合成,其活性受光照强度影响,不受温度直接调控;电子传递链组分包括质体醌(PQ)、细胞色素b6/f复合体、质体蓝素(PC)等,负责传递电子和质子,构建质子梯度;ADP和Pi是合成ATP的原料,NADP⁺是合成NADPH的原料,二者均来自暗反应的消耗与循环。反应过程:三大关键步骤
光反应的全过程可分为光能吸收与传递、电子传递与水的光解、光合磷酸化三个连续且紧密关联的步骤,三个步骤协同作用,完成光能到化学能的转化,同时产生氧气、ATP和NADPH三大核心产物。整个过程中,电子传递、质子转移和能量转化同步进行,形成一个完整的反应体系。一、光能吸收与传递:原初反应
光能吸收与传递是光反应的起始步骤,又称原初反应,发生在类囊体薄膜的色素蛋白复合体上,核心是“捕获光能并传递至反应中心”。该步骤不需要酶的催化,仅依赖光合色素的电子跃迁,是一个快速的物理化学过程,耗时仅为10⁻¹²~10⁻⁹秒。首先,分布在类囊体薄膜上的天线色素(叶绿素b、类胡萝卜素、藻胆素等)吸收可见光的光子,其电子从基态跃迁到激发态。激发态的电子具有较高的能量,但极不稳定,会迅速通过诱导共振的方式,将能量传递给相邻的天线色素,最终传递至反应中心的叶绿素a分子。在这个过程中,能量不断传递,但电子本身并未移动,仅实现了“能量的传递”,类似于“接力赛”,最终将分散的光能集中到反应中心。反应中心的叶绿素a分子接收能量后,其电子被激发至更高的能级,成为高能电子,此时叶绿素a分子因失去电子而带正电,形成“空穴”,完成光能到电能的初步转化。这一步是原初反应的核心,也是光反应中唯一能将光能转化为电能的步骤,只有少数特殊状态下的叶绿素a分子(如P680、P700)能作为反应中心,承担电子激发和转移的功能——其中P680对应光系统Ⅱ,P700对应光系统Ⅰ,二者协同作用,完成后续的电子传递过程。需要注意的是,原初反应中,天线色素仅负责吸收和传递光能,不能参与电子转移,只有反应中心的叶绿素a能完成电子的激发与释放;若光照强度不足,天线色素吸收的光能无法有效传递至反应中心,会导致光反应速率下降。二、电子传递与水的光解
电子传递与水的光解是光反应的核心步骤,发生在类囊体薄膜上,核心是“利用高能电子构建质子梯度,同时补充电子并产生氧气”。该步骤依赖光系统Ⅰ(PSI)、光系统Ⅱ(PSⅡ)和电子传递链的协同作用,水的光解为电子传递提供源源不断的电子,电子传递过程中伴随质子转移,为后续ATP的合成奠定基础。首先,光系统Ⅱ(PSⅡ)的反应中心叶绿素a(P680)被激发后,释放出高能电子,该电子被初级电子受体(去镁叶绿素)捕获,随后通过电子传递链依次传递给质体醌(PQ)、细胞色素b6/f复合体、质体蓝素(PC),最终传递至光系统Ⅰ(PSI)的反应中心叶绿素a(P700)。在电子传递过程中,高能电子的能量逐渐降低,部分能量被用于将类囊体基质中的氢离子(H⁺)泵入类囊体腔,使类囊体腔内的H⁺浓度高于基质,形成跨类囊体膜的质子电化学梯度(质子浓度差+电位差),这是光合磷酸化的能量来源。由于光系统Ⅱ的P680释放电子后形成空穴,需要补充电子才能维持反应的持续进行,此时水会在光和水裂解酶的作用下发生光解反应(又称水的光解)。水的光解反应式为:2H₂O → 4H⁺ + O₂↑ + 4e⁻,即一分子水被分解为两分子氢离子、一分子氧气和四分子电子,其中电子用于补充P680的空穴,使叶绿素a分子恢复基态,继续吸收光能;氢离子一部分进入类囊体腔,增强质子梯度,另一部分用于后续NADPH的合成;氧气则作为副产物释放到细胞外,成为地球上氧气的主要来源。光系统Ⅰ(PSI)的P700接收来自电子传递链的电子后,被光能再次激发,释放出更高能量的电子,该电子被电子受体捕获后,最终传递给NADP⁺,在NADP⁺还原酶的催化下,与类囊体基质中的H⁺结合,形成还原型辅酶Ⅱ(NADPH)。NADPH的形成过程可表示为:NADP⁺ + 2e⁻ + H⁺ → NADPH,NADPH中储存着活跃的化学能,同时具有还原性,将作为暗反应的还原剂,参与二氧化碳的固定与还原。电子传递过程分为非循环电子传递和循环电子传递两种类型:非循环电子传递是主流形式,即电子从水出发,经过光系统Ⅱ、电子传递链、光系统Ⅰ,最终传递至NADP⁺,全程电子不循环,同时产生O₂、ATP和NADPH;循环电子传递仅发生在光系统Ⅰ,电子传递过程中不涉及水的光解,不产生O₂和NADPH,仅产生ATP,用于补充暗反应中ATP的不足,当NADPH积累过多时,循环电子传递会被激活,调节光反应产物的比例。三、光合磷酸化:ATP的合成
光合磷酸化是光反应的最后一步,核心是“利用质子梯度的能量,将ADP和Pi合成ATP”,发生在类囊体薄膜上,依赖ATP合成酶的催化作用,是光能转化为化学能的关键环节——将质子梯度的势能转化为ATP中活跃的化学能,为暗反应提供能量。在电子传递过程中,类囊体腔内的H⁺浓度远高于基质,形成了跨膜的质子电化学梯度,H⁺会顺着浓度梯度,通过类囊体膜上的ATP合成酶(一种通道蛋白)从类囊体腔流向基质。H⁺的流动会驱动ATP合成酶的构象发生变化,这种构象变化会催化ADP和Pi发生脱水缩合反应,合成ATP(三磷酸腺苷),反应式为:ADP + Pi + 能量(来自质子梯度) → ATP。ATP合成酶的结构特殊,分为头部、柄部和基部三部分,基部镶嵌在类囊体薄膜上,形成H⁺通道,头部位于类囊体基质一侧,负责催化ATP的合成。当H⁺通过通道流动时,会带动头部的亚基旋转,使ADP和Pi结合形成ATP,实现“势能→化学能”的转化。这种利用光能驱动质子梯度,进而合成ATP的过程,称为光合磷酸化,与细胞呼吸中的氧化磷酸化(利用化学能驱动质子梯度合成ATP)原理相似,但能量来源不同。光合磷酸化的效率受质子梯度大小的影响,而质子梯度的大小又取决于电子传递的速率和水的光解速率:光照强度越强,电子传递速率越快,水的光解越旺盛,质子梯度越大,ATP的合成效率越高;反之,光照强度不足,质子梯度减小,ATP的合成效率会下降。此外,ATP合成酶的活性也会影响光合磷酸化的效率,其活性受光照调控,黑暗条件下会被抑制,确保光反应仅在光照下进行。至此,光反应的三个步骤全部完成,最终产生三大产物:氧气(副产物,释放到外界)、ATP(能量载体,用于暗反应)、NADPH(还原剂,用于暗反应),同时完成了光能→电能→活跃化学能的转化,为暗反应的顺利进行提供了必要的物质和能量支持。
")
反应特点与影响因素
一、核心反应特点
光合作用光反应阶段具有明确的特异性和规律性,其核心特点可总结为以下四点,与暗反应形成鲜明对比,也是区分两个阶段的关键依据。第一,必须依赖光照:光反应的起始步骤(光能吸收与传递)需要光照驱动,没有光照,光合色素无法吸收光能,电子无法被激发,整个光反应过程会立即停止。即使有充足的原料(水、ADP、Pi等)和酶,黑暗条件下也无法进行光反应,但少数光合细菌可利用化学能替代光能,进行类似光反应的过程,属于特殊情况。第二,反应场所固定:真核生物的光反应仅发生在叶绿体类囊体薄膜上,原核生物仅发生在细胞质膜上,这是因为光合色素、电子传递链组分和ATP合成酶均固定在这些膜结构上,膜结构的分隔是形成质子梯度、实现能量转化的必要条件,脱离膜结构,光反应无法正常进行。第三,与温度无直接关联:光反应主要是光化学反应和物理过程,依赖光合色素和电子传递链的作用,酶仅起到辅助催化作用(如水裂解酶、ATP合成酶),且这些酶的活性受光照调控,不受温度直接影响。因此,在一定温度范围内(如0~30℃),温度变化对光反应速率的影响极小,这与暗反应(酶促反应,受温度影响显著)有本质区别。第四,产物具有瞬时性:光反应产生的ATP和NADPH均为活跃的化学能载体,不稳定,无法长期储存,会迅速被暗反应消耗,若暗反应受阻,ATP和NADPH会积累,进而抑制光反应的进行;而氧气作为副产物,会持续释放到外界,成为大气中氧气的主要来源。二、主要影响因素
光反应的速率受多种因素影响,核心是影响光能吸收、电子传递或质子梯度形成的因素,主要包括光照强度、光合色素含量、水分供应、矿质元素等,各因素相互作用,共同调控光反应的效率。1. 光照强度:这是影响光反应速率的最核心因素。在一定范围内,光照强度越强,光合色素吸收的光能越多,电子激发和传递的速率越快,水的光解和ATP、NADPH的合成效率越高,光反应速率随之提升;当光照强度达到饱和点后,光合色素已达到最大光能吸收能力,电子传递链达到最大负荷,此时光照强度继续增加,光反应速率不再提升,甚至会因强光导致光合色素被破坏(光抑制),使光反应速率下降。2. 光合色素含量:光合色素是捕获光能的核心,其含量直接影响光能吸收效率。叶片中光合色素含量越高,吸收的光能越多,光反应速率越快;反之,光合色素含量不足(如叶片黄化、衰老),会导致光能吸收不足,光反应速率下降。影响光合色素含量的因素包括光照条件(弱光下叶绿素合成减少)、矿质元素(如镁是叶绿素的组成成分,缺镁会导致叶绿素合成不足)、温度(极端温度会破坏叶绿素)等。3. 水分供应:水是光反应的原料,同时也是电子供体,水分不足会直接影响水的光解,导致电子供应不足,电子传递链受阻,进而影响ATP和NADPH的合成;此外,水分不足会导致植物叶片气孔关闭,间接影响暗反应,但对光反应的影响主要体现在原料供应上。4. 矿质元素:多种矿质元素参与光反应的核心过程,如镁(叶绿素的组成成分)、铁(电子传递链中细胞色素的组成成分)、锰(水裂解酶的组成成分)、磷(ATP和NADP⁺的组成成分)等。缺乏这些矿质元素,会导致光合色素合成不足、电子传递受阻或ATP合成障碍,进而降低光反应速率。5. 光照波长:光合色素对不同波长的可见光吸收效率不同,叶绿素主要吸收蓝紫光和红光,类胡萝卜素主要吸收蓝紫光,因此,光照波长会影响光能吸收效率,进而影响光反应速率。例如,红光和蓝紫光照射下,光反应速率较高,而绿光照射下,光反应速率较低(叶绿素对绿光吸收最少)。与暗反应的区别与联系
光合作用的光反应和暗反应(碳反应)是两个相互独立、又相互依存的阶段,二者在反应场所、条件、过程、产物等方面存在明显区别,但共同完成光能到化学能的转化,以及有机物的合成,二者的区别与联系如下:一、核心区别
1. 反应场所不同:光反应发生在类囊体薄膜(真核生物)或细胞质膜(原核生物);暗反应发生在叶绿体基质(真核生物)或细胞质(原核生物),不依赖膜结构。2. 反应条件不同:光反应必须依赖光照,需要光合色素、光反应酶、水;暗反应不需要光照(可在黑暗中进行),需要暗反应酶、ATP、NADPH、二氧化碳(CO₂)。3. 能量转化不同:光反应的能量转化是“光能→电能→活跃的化学能(ATP、NADPH)”;暗反应的能量转化是“活跃的化学能(ATP、NADPH)→稳定的化学能(糖类等有机物)”。4. 核心过程不同:光反应的核心是光能吸收、电子传递、水的光解和光合磷酸化;暗反应的核心是二氧化碳的固定和C3的还原。5. 产物不同:光反应的产物是O₂、ATP、NADPH;暗反应的产物是糖类(如葡萄糖)、ADP、Pi、NADP⁺(可循环用于光反应)。6. 受温度影响不同:光反应与温度无直接关联,受光照强度影响显著;暗反应是酶促反应,受温度影响显著,温度过高或过低都会抑制暗反应酶的活性。二、密切联系
光反应和暗反应相互依存、相互制约,二者共同构成完整的光合作用过程,缺一不可。一方面,光反应为暗反应提供“能量和还原剂”:暗反应中二氧化碳的固定和C3的还原需要能量(由ATP提供)和还原剂(由NADPH提供),若光反应受阻,ATP和NADPH无法合成,暗反应会立即停止;即使有充足的CO₂和暗反应酶,没有光反应提供的物质和能量,暗反应也无法进行。另一方面,暗反应为光反应提供“循环原料”:暗反应消耗ATP和NADPH,产生ADP、Pi和NADP⁺,这些物质会被光反应重新利用,用于合成新的ATP和NADPH;若暗反应受阻,ADP、Pi和NADP⁺无法再生,会导致光反应产生的ATP和NADPH积累,进而抑制光反应的电子传递和光合磷酸化,使光反应速率下降。此外,光反应和暗反应的速率相互协调,当光照强度增加时,光反应速率提升,产生更多的ATP和NADPH,会促进暗反应速率提升;当CO₂浓度增加时,暗反应速率提升,消耗更多的ATP和NADPH,会促进光反应速率提升,二者形成动态平衡,确保光合作用的高效进行。生理意义与应用价值
一、生理意义
光合作用光反应阶段是地球上生命活动的基础,其生理意义不仅体现在光合作用本身,更对整个地球生态系统和生命演化具有重要作用,主要体现在以下三个方面。1. 为地球提供氧气,维持碳氧平衡:光反应中,水的光解产生氧气,这是地球上氧气的主要来源(约占大气中氧气的90%以上)。氧气的产生为需氧生物(如动物、人类)的生存提供了必要条件,同时,氧气参与地球的碳氧循环——光反应释放的氧气,被需氧生物用于呼吸作用,产生CO₂,而CO₂又被植物用于光合作用,形成循环,维持了大气中O₂和CO₂的相对稳定,避免了CO₂过度积累导致的温室效应,也避免了O₂不足导致的需氧生物灭绝。2. 实现光能转化,为生物界提供能量:光反应将太阳能转化为ATP和NADPH中活跃的化学能,随后通过暗反应转化为糖类等有机物中稳定的化学能,这些化学能是整个生物界能量的来源。植物通过光合作用合成有机物,不仅为自身生长发育提供能量,还为食草动物、食肉动物等消费者提供食物和能量,形成食物链和食物网,支撑整个生态系统的运转;此外,煤炭、石油等化石燃料,其本质是古代植物光合作用储存的能量,也是人类社会的主要能源来源。3. 为暗反应提供物质基础,推动有机物合成:光反应产生的ATP和NADPH,是暗反应顺利进行的必要条件,没有光反应,暗反应无法合成糖类等有机物,植物无法生长发育,整个生物界的物质循环也会中断。同时,光反应中产生的氢离子和电子,参与NADPH的合成,为暗反应中CO₂的还原提供了还原剂,推动了无机碳(CO₂)向有机碳(糖类)的转化,实现了碳元素的循环。
")
二、应用价值
基于光反应的原理和特点,人类在农业生产、新能源开发等领域进行了广泛应用,核心是通过调控光反应速率,提高光合作用效率,或模拟光反应过程,开发新型能源技术。1. 农业生产中的应用:在农业生产中,通过改善光照条件、补充矿质元素、合理灌溉等方式,提高光反应速率,进而提高农作物的光合作用效率,增加产量。例如,合理密植可提高田间光照利用率,让更多的叶片吸收光能,提升光反应速率;补充镁肥,促进叶绿素合成,增强光能捕获能力;合理灌溉,保证水的供应,避免因水分不足导致光反应受阻。此外,培育高光效作物品种(如叶绿素含量高、光反应酶活性强的品种),也是提高光反应效率、增加产量的重要途径。2. 新能源开发中的应用:模拟光反应的过程,开发人工光合系统,将太阳能转化为化学能(如氢气、甲醇等),是新能源开发的重要方向。光反应中,水的光解产生氢气(部分光合细菌的光反应可直接产生氢气),人工光合系统通过模拟光合色素的光能吸收、电子传递和水的光解过程,实现水的光解产氢,为清洁能源开发提供了新的思路;此外,通过模拟光合磷酸化过程,开发太阳能电池,将光能直接转化为电能,也具有广阔的应用前景。3. 环境治理中的应用:利用光合生物的光反应,处理污水中的有害物质(如重金属离子、有机物),实现环境治理。例如,蓝细菌的光反应可吸收污水中的重金属离子,将其转化为无害物质;同时,蓝细菌通过光反应释放氧气,可增加污水中的溶解氧,促进污水中有机物的分解,改善水质。此外,大规模种植光合植物(如藻类、芦苇),可吸收大气中的CO₂,缓解温室效应,改善生态环境。深度解读
光合作用光反应阶段看似是一个简单的光化学反应过程,但其本质是地球生命与太阳能相互作用的核心机制,是地球生态系统得以稳定运转的基础,其深层意义远超“产生氧气、合成ATP”的表面功能,可从三个维度进行深度解读。从生命演化的角度来看,光反应的出现是地球生命演化的关键转折点。在地球早期,大气中没有氧气,只有厌氧生物生存,而光反应的出现(最早出现在蓝细菌中),使得光合生物能够利用太阳能分解水,产生氧气,逐渐改变了地球的大气成分,为需氧生物的出现和演化提供了条件。同时,光反应实现了“光能→化学能”的转化,让生物摆脱了对化学能(如有机物分解)的依赖,拓宽了生命的生存范围,推动了生物的多样化演化——从简单的光合细菌,到复杂的绿色植物,光反应的机制不断完善,成为生命演化的重要驱动力。从能量循环的角度来看,光反应是地球上能量循环的起点。太阳能是地球上最丰富的清洁能源,光反应将太阳能转化为化学能,储存于ATP和NADPH中,再通过暗反应转化为有机物中的稳定化学能,这些化学能通过食物链和食物网传递给整个生物界,最终通过呼吸作用分解,释放出能量,同时产生CO₂和水,重新用于光合作用,形成“太阳能→化学能→热能”的循环。这种能量循环,不仅支撑着生物界的生命活动,也维持着地球的生态平衡,是地球区别于其他行星的重要特征。从分子机制的角度来看,光反应是一个高度精密、协同的分子体系。光合色素的光能吸收与传递、电子传递链的电子转移、质子梯度的构建、ATP和NADPH的合成,每一个步骤都依赖于特定的分子(色素、酶、电子载体)协同作用,形成一个高效、有序的反应体系。这个体系的精密性,体现了生物在长期演化过程中的适应性,也为人类模拟光合过程、开发新能源提供了灵感——例如,人工光合系统的研发,就是基于光反应的分子机制,模拟光合色素的光能捕获、电子传递和水的光解过程,实现太阳能的高效转化。此外,光反应的调控机制也具有重要的研究价值。光反应的速率受光照强度、光合色素含量等因素的调控,同时也受暗反应的反馈调节,这种调控机制确保了光合作用的高效、稳定进行,避免了能量的浪费和物质的积累。深入研究光反应的调控机制,不仅有助于我们理解光合作用的本质,还能为农业生产、新能源开发提供更精准的理论指导,例如,通过调控光反应的关键酶活性,提高农作物的光合效率,或优化人工光合系统的性能,提升太阳能转化效率。最新消息
近年来,随着分子生物学、纳米技术等学科的发展,关于光合作用光反应阶段的研究取得了多项重要突破,不仅深化了人类对光反应机制的理解,也为人工光合技术的开发提供了新的思路,以下是近期的核心研究进展。
2026年2月26日,中国科学院上海有机化学研究所等研究团队在人工光合组装体研究中取得重大进展,成功构建了一类新型球形色素体纳米胶束,实现了紫细菌光捕获复合物(LH2与LH1–RC)超级复合物结构模拟及水相高效光催化制氢,该系统的催化转化数(TON)突破了667000,为模拟光反应机制、开发高效人工光合系统提供了新路径。该研究团队受天然光合系统的启发,发现光合生物在长期进化过程中演化出近乎完美的光捕获系统,其中LH2和LH1–RC通过环状蛋白骨架对细菌叶绿素分子的精准排布,构筑出环形阵列结构,量子效率可达95%以上。基于这一结构特征,研究团队设计了两亲性卟啉基细菌叶绿素类似物(BChlA)分子,通过无蛋白骨架条件下的自组装,构建了超分子阴离子型卟啉组装体(SAPA),并与带正电的镍基分子催化剂通过分级共组装精准结合,成功模拟了天然色素体“天线—反应中心”关键超级复合物结构——这一结构与光反应中“天线色素捕获光能、反应中心转化光能”的核心机制高度一致。
研究团队通过冷冻电镜技术,首次直接观测到仿生体系中卟啉环状阵列形成的高分辨关键证据,在纳米胶束表面清晰分辨出约22个环状亚基,每个环由12个卟啉分子组成,且纳米胶束表面通过超分子多价性耦合催化反应中心,形成了类似天然LH1–RC结构的核心反应区。稳态吸收光谱研究表明,催化剂与卟啉阵列结合后,体系的Soret吸收峰从424纳米红移至436纳米,Q带同步位移,呈现出与天然色素体中B850向B875类似的吸收和能量传递路径转变,进一步验证了该仿生系统对天然光反应机制的模拟效果。在光催化性能方面,该人工色素体系统在水相可见光驱动下,表现出优异的催化产氢活性与稳定性。在可见光(AM1.5G,420Cut-off Filter)照射下,该组装体体系的产氢效率较非组装自由分子体系提升了40倍;连续运行72小时后,催化位点TON达到667321,24小时内测定的外量子效率为6.8%(435纳米)。飞秒瞬态吸收光谱研究进一步揭示了其高效催化的微观动力学机制:组装体中光生电子由卟啉三线态向催化剂的转移时间缩短至皮秒量级,电荷分离态寿命延长至1.35纳秒,形成“快速转移、缓慢复合”的电荷分离态,有效抑制了能量耗散,这与天然光反应中电子传递的高效性机制高度契合。
该研究的突破,不仅成功模拟了天然光合系统光反应阶段的核心结构和功能特征,还发展了基于超分子组装策略构建水相高效人工光合产氢体系的新途径,为太阳能燃料合成系统的设计和机制解析提供了重要的结构基础与思路。未来,随着研究的深入,这类人工光合系统有望实现规模化应用,为清洁能源开发、碳达峰碳中和目标的实现提供重要支撑。此外,近期还有研究团队通过基因编辑技术,优化了植物光反应中关键酶(如ATP合成酶、水裂解酶)的活性,成功提高了植物的光反应速率和光合效率,为培育高光效作物品种提供了新的技术手段。该研究发现,通过编辑ATP合成酶的相关基因,可增强质子梯度的利用效率,提高ATP的合成速率,同时促进NADPH的生成,进而推动暗反应的进行,最终提高农作物的产量和品质,目前该技术已在水稻、小麦等作物中进行了初步试验,取得了良好的效果。[1][2]
词条图片